
云南虫化石
(神秘的地球uux.cn报导)据知识分子(撰文 姜宝玉):编者按:7月25日,中国科学院院士、西北大学教授舒德干向《知识分子》投稿,就7月8日Science杂志发表的一项探究距今5.18亿年的澄江生物群云南虫的岗位,提出了各异看法(争鸣 | 舒德干:被热炒的云南虫,究竟是沈阳裁员消息热点不是脊椎动物?)。对此,该论文的联合通讯作者、南京大学地球科学与工程学院教授姜宝玉近期来稿,对舒文做了答复。
《知识分子》于7月25日发表了《争鸣|舒德干:被热炒的云南虫,究竟是不是脊椎动物?》一文。文中,西北大学舒德干院士对我们2022年7月8日发表于Science的论文[1](以下简称 “论文”)开展了评述(以下简称 “评述”)。如今,由我代表探究团队对 “评述” 中的要点做简易答复。
先是,致谢 “评述” 对 “论文” 表达了肯定和鼓励。“评述” 中强调,“文章面世后,被境内各大传媒炒得沸沸扬扬,给学界和大众导致了显著的误导”。一些传媒转载我们官网资讯,一些传媒转载境外的资讯或Science同期发表的评述文章,乃至含有一些访谈过我们的资讯中,或许使用了一些不够严谨的表达方式,对此我们表达歉意。另外,“评述” 也从论文 “微观结构的多解性” “论证的逻辑性” 和 “证据的科学性” 三方面对论文提出了质疑。由于 “评述” 中没有对 “微观结构的多解性” 展开研究,我这里仅就后两方面的质疑做些说明,供舒院士和广大读者研究。
一些基础概念
由于 “论文” 的核心信息之一是研究云南虫在生命树上的操控系统发生位置,在说明之前,我先简易说明生物操控系统发生关系(生命树)中关乎的一些基础概念,便于大家理解。
操控系统发生框架反映了生物类群之间的亲缘关系和生物演化的过程,是现代演化生物学普遍使用的自然分类体系 [2]。在对重大生物类群起源和早期演化的探究中,一个重大难题是推测现生物种的最后共同祖先(Last Common Ancestor,LCA或Most Recent Common Ancestor,MRCA)具有哪些性状特征。以便便捷研究生物类群起源和早期演化,演化生物学家提出了冠群、干群和总群等概念 [3,4]。
冠群(crown group)指某一生物类群所有现生物种的最后共同祖先和该祖先形成的所有后代,含有所有现存的和已灭绝的后代(图1)。某一冠群和与它演化关系最近的冠群也存在一个最后共同祖先,而由这一祖先分异出的两个分支则被称为这两个冠群分别对应的总群(total group)。总群中除去冠群物种之后,早于冠群分异的、已灭绝的物种,就是相应冠群对应的干群(stem group)。大家在一些生物演化科普中目睹某某类群的祖先类群通常指某一冠群的干群。
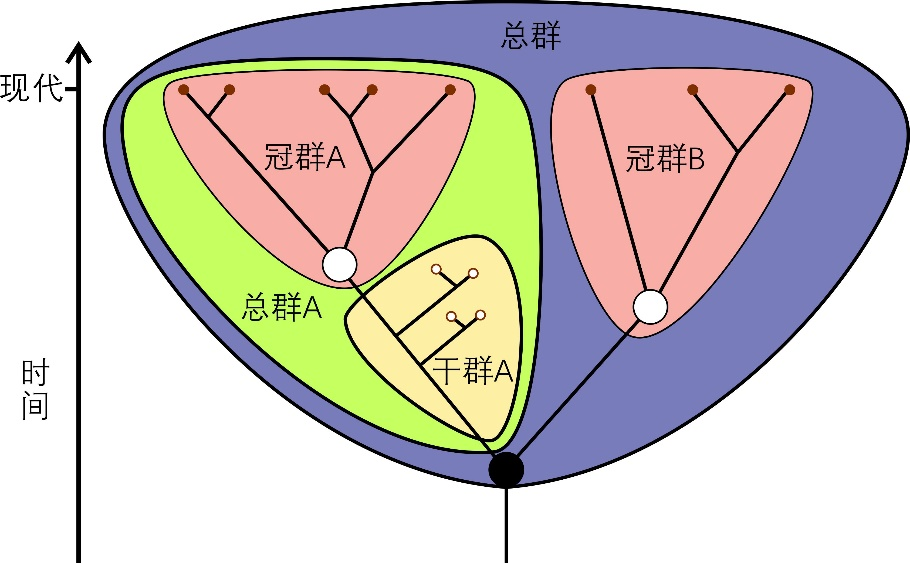
图1 冠群、干群和总群的示意图。图中展示A和B两个冠群、冠群A对应的干群和总群、以及A和B两个冠群构成的总群。红点强调现生类群,小白点强调化石类群,大白点强调相应冠群的最后共同祖先,大黑点强调两个冠群的最后共同祖先 | 更改自维基百科使用者Mattbierner和Peter coxhead(https://en.wikipedia.org/wiki/Crown_group)(CC BY-SA 4.0)。
论证的科学逻辑
“评述” 依据 “任何较高等的类群起源,都应该是由一系列较低等类群,也就是它们的祖先类群,是历程了连续的基因革新,以及由基因革新引发的有关器官构造革新,才一步一步进化来的” 这一生物进化基础逻辑,强调 “绝不能由于某些低等类群,为后代的高等类群创造了某些基因和基础器官,就得出结论说 “前者就是后者”,或者说 “低等动物就是高等动物”。
以便说明这种逻辑关系,“评述” 中列举了三个例子。第一个例子觉得两栖类的一支演化出爬行动物,而爬行动物的一支演化出鸟类。第二个例子提出 “3000万年至5000万年过去,是由某一类相当原始的小猴子” 演化出古猿,而古猿演化出人类。第三个例子列举了半索动物演化出脊索动物,脊索动物演化出脊椎动物。“评述” 中刻画的这种简易化的演化关系是不严谨的,不符合操控系统发生确认反映出的操控系统发生关系。以与“论文”有关的半索动物为例,半索动物在后口动物操控系统树中早已偏离了向脊椎动物冠群演化的支系(图2)。也就是说,半索动物无法演化出脊索动物和脊椎动物,脊索动物是由脊索动物和半索动物的最后共同祖先演化而来。与此相似,两栖类早已偏离向羊膜动物冠群演化的路径,无法演化出爬行动物和鸟类。一文读懂演员阵容资讯另外,生物的演化也不存在从低级到高级这样的演化顺序。
在这三个例子中,“评述” 觉得不能将两栖类称为鸟类,不能将猴子称为人类,也不能将半索动物称为脊椎动物。“评述” 将上述三个例子列举的逻辑关系与 “论文” 联系起来,是由于 “评述” 觉得,“云南虫及其同类进化到脊椎动物的过程是:先由低等的 ‘无脊索动物’,如云南虫,半索动物门,古虫动物门等门类中的某一支,先是进化成具有脊索的 ‘低等脊索动物’(含有头索动物,尾索动物)。再持续进化,便创造出头脑,创造出脊椎,最后变成脊椎动物。这个基础演进路径科学家搞得相当清楚了。” 由于这里没有列参考文献,不清楚这里觉得这种基础演进路径搞得相当清楚的科学家是指哪些学者。有关古虫动物(vetulicolians)和云南虫(yunnanozoans)与现生半索动物、脊索动物和脊椎动物之间的操控系统发生关系,在绝大多数新近的、有关脊椎动物起源的文献中,都被觉得有风波的 [5–7],这一点在 “论文” 的引言若干已有说明:云南虫在各异探究中被置于脊椎动物干群 [8,9]、头索动物 [10]、半索动物 [11]、后口动物干群 [12]、乃至原始的两侧对称动物 [13]。
生物演化关乎众多类群和性状的繁琐过程,无法简易地用从低级到高级的“进化”来刻画。操控系统发生确认就是综合各异生物类群的众多性状特征资料,使用适合的使用和算力,反演重建生物演化的过程。“论文” 的操控系统发生确认使用了舒院士等2017年发表的两篇论文[14,15]中的有关性状资料和有关确认方法,并在此基础上充分考虑了当前探究中化石解剖学构造阐释上的不确定性和风波,采取了保守的性状编码方式(见 “论文” 发表的附件表格),运用当前探究中常用的以概率模型为基础的贝叶斯推断法,得出了云南虫最或许位于图2所示的演化位置。
自然,假如对解剖学性状使用各异观点的阐释和资料编码,或者使用各异的操控系统发生确认方法和模型 [16,17],或许会得出与 “论文” 结局有差异的操控系统发生关系,这是较为和论证各异演化假设中普遍的现象。
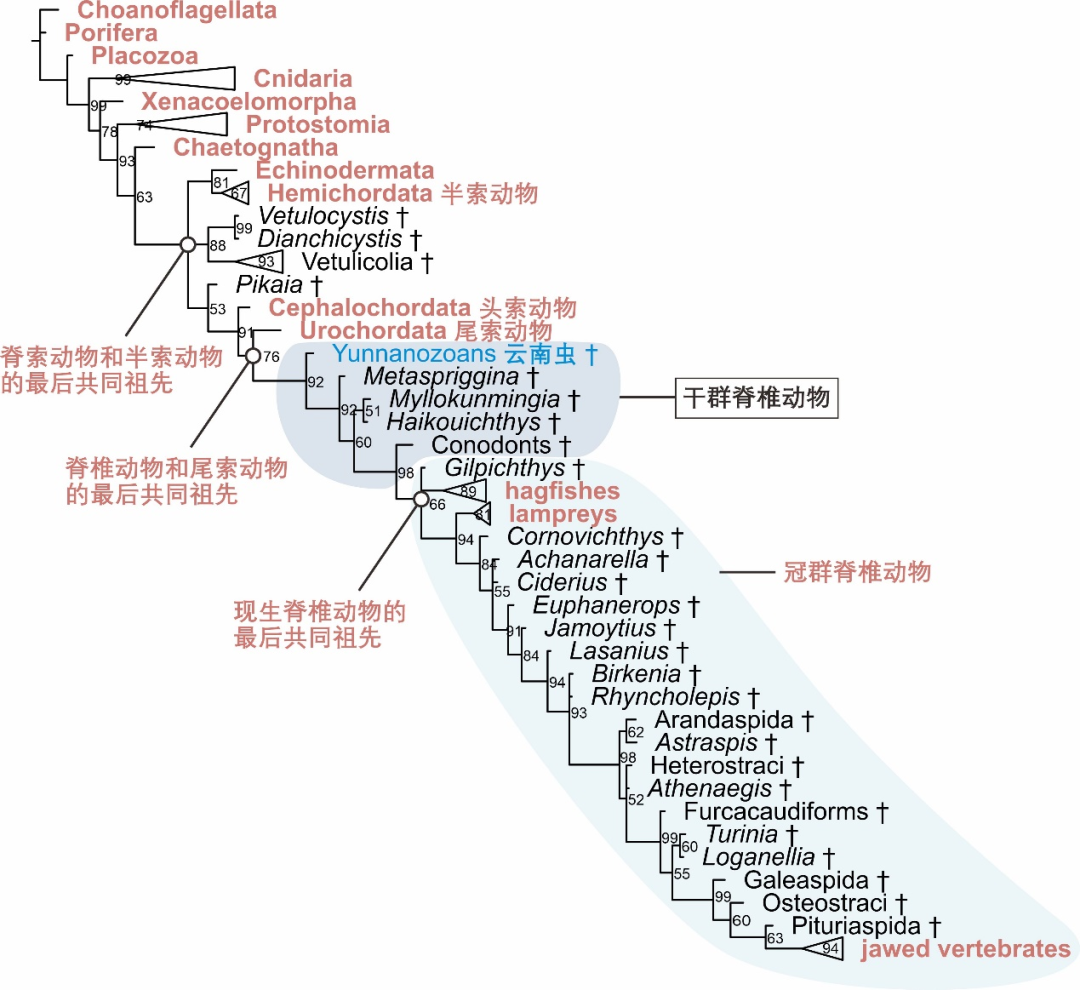
图2 “论文” 中得到的操控系统发生树。现生类群显示为红色,灭绝类群用剑标(†)强调,若干类群合并强调。节点旁的数字强调后验概率(百分比强调)。
脊索动物含有头索动物、尾索动物和脊椎动物。分子操控系统发生探究表明,脊椎动物和尾索动物之间的关系最近,而头索动物归于脊索动物中最初分异出来的一支 [18]。将前面谈及的冠群、干群的概念使用到脊椎动物起源的难题中,可以看出脊椎动物的冠群含有现生的圆口纲和有颌类。而脊椎动物的干群,则含有在脊椎动物总群支系上、位于脊椎动物和尾索动物最后共同祖先和冠群脊椎动物之间的所有已绝灭类群(图2)。“论文” 经由操控系统发生确认揭示,云南虫类的演化位置处于脊椎动物的干群之中,并且是当下已知分异最初的脊椎动物干群支系(图2)。“论文” 从未将半索动物、头索动物或尾索动物称为脊椎动物。
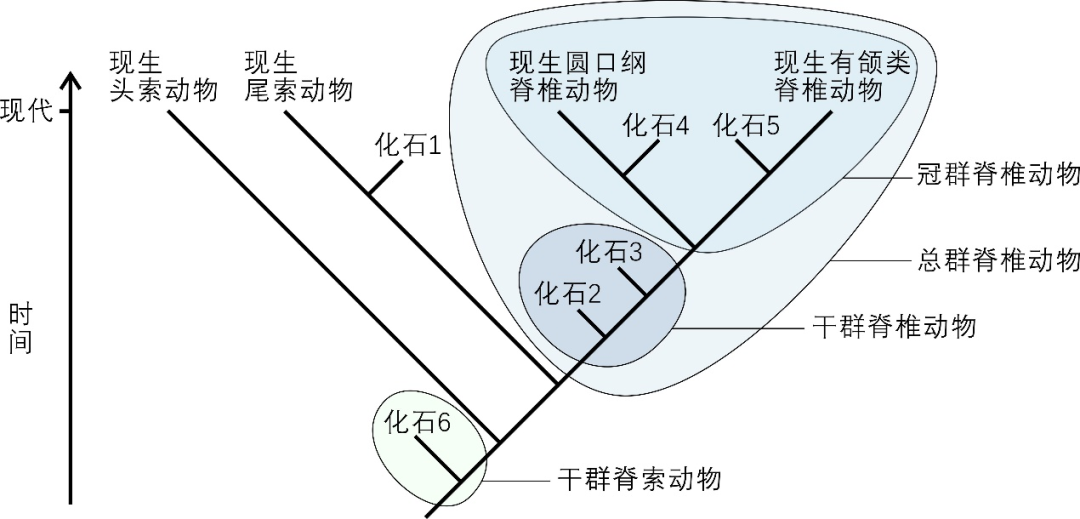
图3 脊索动物操控系统发生树示意图 | 依据文献[4]绘制
证据的科学性
“评述” 觉得,“云南虫除了具有鳃裂外,不具备其余核心的五个革新特征,即没有脊索,没有肌节,没有肛后尾,没有脊椎骨,也没有头脑眼。所以,云南虫与脊椎动物毫不相干系。”
先是,许多重大生物类群的起源与演化探究表明,冠群拥有的一系列性状特征往往是在该类群起源和早期演化中逐步获得的 [19]。某一类群的干群物种,往往只具有其冠群的少数性状特征,代表了演化过程中的过渡类型。云南虫不具有冠群脊椎动物的一些性状特征,也无法否定云南虫归于干群脊椎动物。
其次,化石曾经是活着的生物。生物从死亡、腐烂到被掩埋,最后变成化石,会历程漫长的详细票房排行榜单地质时间和繁琐的生物、化学和物理转变 [20]。在这个过程中,很多生物特征会消失,或者留下的证据没有被观察到,所以化石保存的特征远少于现生动物。所以,化石中缺失的特征是无法确定曾经是否存在的,尤其是软组织特征 [5]。云南虫化石也存在相似的状况。云南虫化石的许多宏观结构,各异探究团队有各异的阐释。例如:
1.脊索。脊索动物的脊索是由含有胶原蛋白的外鞘和内部的细胞组成的 [21]。尽管不轻松腐烂 [6],但是或许由于不轻松碳化,脊索在化石中并不普遍 [20]。化石圆口纲脊椎动物希罗卡似盲鳗(Myxinikela siroka)[22,23]、蒙大拿哈迪斯蒂鳗(Hardistiella Montanensis)[24]、詹格尔双鳗(Pipiscius zangerli)[25,26]、乃至许多耳材村海口鱼(Haikouichthys ercaicunensis)标本 [27,28] 都没有观察到脊索。不能由于没有观察到脊索就反驳这些化石是脊索动物。前人也提出了云南虫脊索的阐释 [8–10, 29, 30],尽管存在风波,但是还不能简易地定论云南虫没有脊索。
2.肌节。肌节是分段的躯干肌肉,并不是所有脊索动物都有的特征。头索动物和脊椎动物拥有形态各异的肌节,而尾索动物没有肌节 [31]。所以,不能依据没有肌节来断定云南虫不是脊索动物。云南虫背部的分节被前人阐释为肌节,也察觉有肌肉纤维的构造 [8–10, 29, 30]。尽管存在风波,但是不能轻易反驳已有的肌节证据。即使前方新的证据证明背若干节是表皮结构,也不能排除云南虫在表皮之下肌节曾经存在的或许性。
3.肛后尾。云南虫的躯干后面存在显著的突出物 [8,9,32]。该结构被前人阐释为肛后尾 [8,9],仅有少数学者对该阐释提出了质疑 [33]。
4.脊椎。脊椎是附着在脊索上分节的骨骼或者软骨。所有的圆口纲脊椎动物中,只有现生七鳃鳗、现生的黏盲鳗属(Eptatretus)的尾部 [34]和有风波的化石圆口纲寻常塔利怪鳗(Tullimonstrum gregarium)[35] 有脊椎,现生的盲鳗属(Myxine)[36] 和其他所有圆口纲化石都没有察觉脊椎。不能由于这些脊椎动物没有脊椎就说它们不归于脊椎动物。
5.头脑眼。有关头,依据拉马克(Lamarck, 1801)[37]的定义,脊椎动物同义词是有头类(Craniota Haeckel, 1866或Craniata Lankester, 1877)[38, 39],意味着它们都有骨质或者软骨的头颅。头颅含有包围大脑的脑颅和包围咽腔的咽颅。无脊椎动物中的头索动物和半索动物也有支撑咽腔的鳃部结构,但都不是由软骨组成的。“论文” 提供了云南虫具有咽部软骨的证据,说明云南虫具有了由软骨支撑的咽腔——原始咽颅。依据传统上脊椎动物的定义,云南虫已然具有了核心的脊椎动物特征。
大脑首要由柔软的神经组织组成,另外也是腐烂最快的器官之一 [6],大多数脊椎动物化石都没有保存大脑。有关眼睛,脊索动物有很各式眼睛,头索动物和尾索动物身体前端都具有微小的感光器官,大若干脊椎动物则具有摄像机型眼 [40]。但是,很多脊椎动物也不具有显著的眼睛。比如最初分化出来的现生脊椎动物盲鳗,它们的眼睛没有晶状体和黑色素,完全被皮肤覆盖 [41]。化石长吻特提斯盲鳗(Tethymyxine tapirostrum)也没有观察到眼睛 [42]。事实上,前人曾经报导过云南虫标本上具有眼睛构造 [8,9,30],只是保存眼睛的标本很少,才存在风波。假如云南虫的眼睛或者感光器官像盲鳗或者头索动物那样不显著,很难在化石中保存或找到眼睛的痕迹。
另外,“评述” 后记中谈及,“新文章将云南虫归入脊椎动物的首要微观构造证据是,它与后者具有相似的 ‘细胞软骨’”,“该性状具有多解性和不确定性,很或许广泛呈现于众多无脊椎动物类群”。“论文” 并没有反驳无脊椎动物也有软骨,文中也举出了头索动物口须软骨的例子 [1]。“论文” 提供的首要微观证据不是云南虫具有细胞软骨,而是云南虫具有了脊椎动物特有的特征——鳃弓软骨,这也是 “论文” 的核心证据。
科学探究需要不断探索
我相当致谢舒院士亲自对 “论文” 中和传媒宣传中存在的详细网大电影消息难题提出指责,也使用对舒院士提出的一些质疑表达了各异观点。我身边几代令人尊敬的科学家一直用身体力行的方式教学我,要严谨地对待科学难题,大胆假设,小心求证,我谨记于心。科学探究需要不断探索,身为一支相对年轻的科研团队,我们使用使用多学科交叉的方法,为解决持久以来存在的风波难题,提供一些形态学和超微结构方面的证据。在这一过程中,我们肯定存在这样或者那样的不足。
“论文” 的结论仅代表我们的观点,一定有很多学者持有各异观点。期盼 “论文” 能起到抛砖引玉的效果,吸引同行们在学术领域发表各自的观点。这样前方会有更多的同行留意这一充满魅力的领域,一定会察觉更多的新证据。新证据也许证明我们的结论是不精确的,但我们还是期盼,我们察觉的这些证据是有价值的。最后,也致谢《知识分子》记者部能给我们这样一个机遇,向舒院士和广大读者表达各异的学术观点。
参考文献:
1. Q. Y. Tian, F. C. Zhao, H. Zeng, M. Y. Zhu, B. Y. Jiang, Ultrastructure reveals ancestral vertebrate pharyngeal skeleton in yunnanozoans. Science. 377, 218–222 (2022). https://doi.org/10.1126/science.abm27082. E. O. Wiley, B. S. Lieberman, Phylogenetics: Theory and Practice of Phylogenetic Systematics, Second Edition (John Wiley & Sons, Inc., Hoboken, NJ, USA, 2011). http://doi.wiley.com/10.1002/97811180178833. G. E. Budd, S. Jensen, A critical reappraisal of the fossil record of the bilaterian phyla. Biol. Rev. 75, 253–295 (2000). https://doi.org/10.1111/j.1469-185X.1999.tb00046.x4. R. P. S. Jefferies, “The origin of chordates-a methodological essay” in The Origin of Major Invertebrate Groups, M. R.House, Ed. (Academic Press, London, 1979), pp. 443–477. https://archive.org/details/originofmajorinv0000unse/page/4435. P. C. J. Donoghue, M. A. Purnell, Distinguishing heat from light in debate over controversial fossils. BioEssays. 31, 178–189 (2009). https://doi.org/10.1002/bies.2008001286. R. S. Sansom, S. E. Gabbott, M. A. Purnell, Non-random decay of chordate characters causes bias in fossil interpretation. Nature. 463, 797–800 (2010). https://doi.org/10.1038/nature087457. P. Janvier, Facts and fancies about early fossil chordates and vertebrates. Nature. 520, 483–489 (2015). https://doi.org/10.1038/nature144378. J. Y. Chen, D. Y. Huang, C. W. Li, An early Cambrian craniate-like chordate. Nature. 402, 518–522 (1999). https://doi.org/10.1038/9900809. J. Mallatt, J. Y. Chen, Fossil sister group of craniates: predicted and found. J. Morphol. 258, 1–31 (2003). https://doi.org/10.1002/jmor.1008110. J. Y. Chen, J. Dzik, G. D. Edgecombe, L. Ramsköld, G. Q. Zhou, A possible Early Cambrian chordate. Nature. 377, 720–722 (1995). https://doi.org/10.1038/377720a011. D. G. Shu, X. L. Zhang, L. Chen, Reinterpretation of Yunnanozoon as the earliest known hemichordate. Nature. 380, 428–430 (1996). https://doi.org/10.1038/380428a012. D. G. Shu, S. Conway Morris, Z. F. Zhang, J. N. Liu, J. Han, L. Chen, X. L. Zhang, K. Yasui, Y. Li, A new species of yunnanozoan with implications for deuterostome evolution. Science. 299, 1380–1384 (2003). https://doi.org/10.1126/science.107984613. R. A. Dewel, Colonial origin for Eumetazoa: major morphological transitions and the origin of bilaterian complexity. J. Morphol. 243, 35–74 (2000). https://doi.org/10.1002/(SICI)1097-4687(200001)243:1%3C35::AID-JMOR3%3E3.0.CO;2-%2314. Q. Ou, J. Han, Z. F. Zhang, D. G. Shu, G. Sun, G. Mayer, Three Cambrian fossils assembled into an extinct body plan of cnidarian affinity. Proc. Natl. Acad. Sci. U.S.A. 114, 8835–8840 (2017). https://doi.org/10.1073/pnas.170165011415. J. Han, S. Conway Morris, Q. Ou, D. G. Shu, H. Huang, Meiofaunal deuterostomes from the basal Cambrian of Shaanxi (China). Nature. 542, 228–231 (2017). https://doi.org/10.1038/nature2107216. M. N. Puttick, J. E. O’Reilly, D. E. Pisani, P. C. J. Donoghue, Probabilistic methods outperform parsimony in the phylogenetic analysis of data simulated without a probabilistic model. Palaeontology. 62, 1–17 (2019). https://doi.org/10.1111/pala.1238817. P. A. Goloboff, A. Torres, J. S. Arias, Weighted parsimony outperforms other methods of phylogenetic inference under models appropriate for morphology. Cladistics. 34, 407–437 (2018). https://doi.org/10.1111/cla.1220518. F. Delsuc, H. Brinkmann, D. Chourrout, H. Philippe, Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature. 439, 965–968 (2006). https://doi.org/10.1038/nature0433619. D. H. Erwin, Novelty and innovation in the history of life. Curr. Biol. 25, R930–R940 (2015). https://doi.org/10.1016/j.cub.2015.08.01920. L. A. Parry, F. Smithwick, K. K. Nordén, E. T. Saitta, J. Lozano-Fernandez, A. R. Tanner, J. B. Caron, G. D. Edgecombe, D. E. G. G. Briggs, J. Vinther, Soft-bodied fossils are not simply rotten carcasses - toward a holistic understanding of exceptional fossil preservation. BioEssays. 40, 1700167 (2017). https://doi.org/10.1002/bies.20170016721. G. Annona, N. D. Holland, S. D’Aniello, Evolution of the notochord. EvoDevo. 6, 1–13 (2015). https://doi.org/10.1186/s13227-015-0025-322. S. E. Gabbott, P. C. J. Donoghue, R. S. Sansom, J. Vinther, A. Dolocan, M. A. Purnell, Pigmented anatomy in Carboniferous cyclostomes and the evolution of the vertebrate eye. Proc. R. Soc. B Biol. Sci. 283, 20161151 (2016). https://doi.org/10.1098/rspb.2016.115123. T. Miyashita, A Paleozoic stem hagfish Myxinikela siroka — revised anatomy and implications for evolution of the living jawless vertebrate lineages. Can. J. Zool. 98, 850–865 (2020). https://doi.org/10.1139/cjz-2020-004624. P. Janvier, R. Lund, Hardistiella Montanensis n. gen. et sp. (Petromyzontida) from the Lower Carboniferous of Montana, with remarks on the affinities of the lampreys. J. Vertebr. Paleontol. 2, 407–413 (1983). https://doi.org/10.1080/02724634.1983.1001194325. D. Bardack, E. S. Richardson, New agnathous fishes from the Pennsylvanian of Illinois. Fieldiana Geol. 33, 489–510 (1977). https://doi.org/10.5962/bhl.title.516726. T. Miyashita, R. W. Gess, K. Tietjen, M. I. Coates, Non-ammocoete larvae of Palaeozoic stem lampreys. Nature. 591, 408–412 (2021). https://doi.org/10.1038/s41586-021-03305-927. D. G. Shu, H. L. Luo, S. Conway Morris, X. L. Zhang, S. X. Hu, L. Chen, J. Han, M. Zhu, Y. Li, L. Z. Chen, Lower Cambrian vertebrates from south China. Nature. 402, 42–46 (1999). https://doi.org/10.1038/4696528. X. G. Zhang, X. G. Hou, Evidence for a single median fin-fold and tail in the Lower Cambrian vertebrate, Haikouichthys ercaicunensis. J. Evol. Biol. 17, 1162–1166 (2004). https://doi.org/10.1111/j.1420-9101.2004.00741.x29. J. Dzik, Yunnanozoon and the ancestry of chordates. Acta Palaeontol. Pol. 40, 341–360 (1995). https://bibliotekanauki.pl/articles/1994630. J. Y. Chen, “The origins and key innovations of vertebrates and arthropods” in Earth and Life (Springer Netherlands, Dordrecht, 2012), pp. 239–379. http://link.springer.com/10.1007/978-90-481-3428-1_1031. T. Onai, The evolutionary origin of chordate segmentation: revisiting the enterocoel theory. Theory Biosci. 137, 1–16 (2018). https://doi.org/10.1007/s12064-018-0260-y32. P. Y. Cong, X. G. Hou, R. J. Aldridge, M. A. Purnell, Y. Z. Li, New data on the palaeobiology of the enigmatic yunnanozoans from the Chengjiang Biota, Lower Cambrian, China. Palaeontology. 58, 45–70 (2015). https://doi.org/10.1111/pala.1211733. D. G. Shu, S. Conway Morris, Response to comment on “A new species of yunnanozoan with implications for deuterostome evolution.” Science. 300, 1372–1372 (2003). https://doi.org/10.1126/science.108557334. K. G. Ota, S. Fujimoto, Y. Oisi, S. Kuratani, Identification of vertebra-like elements and their possible differentiation from sclerotomes in the hagfish. Nat. Commun. 2, 373–376 (2011). https://doi.org/10.1038/ncomms135535. V. E. McCoy, E. E. Saupe, J. C. Lamsdell, L. G. Tarhan, S. McMahon, S. Lidgard, P. Mayer, C. D. Whalen, C. Soriano, L. Finney, S. Vogt, E. G. Clark, R. P. Anderson, H. Petermann, E. R. Locatelli, D. E. G. Briggs, The ‘Tully monster’ is a vertebrate. Nature. 532, 496–499 (2016). https://doi.org/10.1038/nature1699236. K. G. Ota, Y. Oisi, S. Fujimoto, S. Kuratani, The origin of developmental mechanisms underlying vertebral elements: implications from hagfish evo-devo. Zoology. 117, 77–80 (2014). https://doi.org/10.1016/j.zool.2013.10.01037. J.-B. de M. de Lamarck, Système des animaux sans vertèbres ou tableau général des classes, des ordres et des genres de ces animaux (L’auteur, 1801). https://doi.org/10.5962/bhl.title.1425538. E. Haeckel, Generelle Morphologie der Organismen (1866). https://www.degruyter.com/document/doi/10.1515/9783110848281/html?lang=en39. E. R. Lankester, Memoirs: notes on the embryology and classification of the animal kingdom: comprising a revision of speculations relative to the origin and significance of the germ-layers. J. Cell Sci. s2-17, 399–454 (1877). https://doi.org/10.1242/jcs.s2-17.68.39940. S. N. Bradshaw, W. T. Allison, Hagfish to illuminate the developmental and evolutionary origins of the vertebrate retina. Front. Cell Dev. Biol. 10, 1–20 (2022). https://doi.org/10.3389/fcell.2022.82235841. E. M. Dong, W. T. Allison, Vertebrate features revealed in the rudimentary eye of the Pacific hagfish (Eptatretus stoutii). Proc. R. Soc. B Biol. Sci. 288, 1–21 (2021). https://doi.org/10.1098/rspb.2020.218742. T. Miyashita, M. I. Coates, R. Farrar, P. Larson, P. L. Manning, R. A. Wogelius, N. P. Edwards, J. Anné, U. Bergmann, A. R. Palmer, P. J. Currie, Hagfish from the Cretaceous Tethys Sea and a reconciliation of the morphological–molecular conflict in early vertebrate phylogeny. Proc. Natl. Acad. Sci. 116, 2146–2151 (2019). https://doi.org/10.1073/pnas.1814794116
有关报导:争鸣 | 舒德干:被热炒的云南虫,究竟是不是脊椎动物?

云南虫复原图 杨定华
(神秘的地球uux.cn报导)据知识分子(撰文 舒德干):编者按:本年7月8日,《科学》(Science)杂志发表了一项由中国科学院南京地质古生物探究所与南京大学兴办达成的探究成果,并同期配发了一位国外同行撰写的短评。这篇文章首要探究了距今5.18亿年的澄江生物群察觉的云南虫的咽弓具有的超微结构特征,觉得其叠盘状细胞软骨及微原纤维构造主导的细胞间基质为脊椎动物所特有。另外,文章依据新近的操控系统发育确认得出结论,觉得操控系统分类位置持久存在风波的云南虫为脊椎动物最原始的类群。文章发表后,境内外传媒均有众多报导,也引发了有关同行的 “研究”。昨日,西北大学舒德干院士向《知识分子》投稿,就此发表他的看法。
今日很高兴给大家转发一个有趣的议题,它在科学上也是一个很重大的课题。之所以说它重大,是由于它关乎到我们人类,尽管不直接研究现代人类起源,但与我们人类所属的脊椎动物全部大家人起源密切有关。
大家都得知,早在200年前,即1822年,进化论先驱——法国博物学家拉马克(Jean-Baptiste Lamarck),就将全部动物界确认为脊椎动物和无脊椎动物两大若干。从那时以来,脊椎动物是如何从无脊椎动物逐步演化而来,就一直受到学者们的尤其留意。

图1进化论的先驱——法国博物学家拉马克
在今日这个备受留意的题目里,我们重点研究,“云南虫究竟是不是脊椎动物”?
今日谈这个议题不是心血来潮,它事出有因。由于前几天我国有几个学者,在美国的一份著名杂志Science(《科学》)上发表了一篇论文,对云南虫开展了微观形态学的探究,得出了一个颇为轰动的结论—— “云南虫是原始的脊椎动物”!

图2 2022年7月15日出版的《科学》,刊载了有关云南虫的探究论文
这项探究是值得鼓励的,文章也具有一定的科学参考价值。但客观地说,这个探究还不太成熟,证据和结论都存在较为大的难题。尤其是,文章面世后,被境内各大传媒炒得沸沸扬扬,给学界和大众导致了显著的误导,有必要予以公开澄清。
我之所以说这个探究有难题,首要是基于以下几个方面:
第一,它谈到云南虫的这些微观结构具有 “多解性”,也就是具有很大的 “不确定性”,可以有各式阐释。假如有各式阐释,就或许得出 “各式结论”。
第二,文章的逻辑有难题。科学逻辑在我们进化生物学上就是生命是 “逐步进化” 的逻辑,知晓这一点相当重大。
第三,就是 “证据”。事实证据至关重大,依据不可靠的证据,其结论自然不可靠。
我国对云南虫做透彻探究的学术团队首要有四个,除了该文团队外,其余三个团队都觉得 “云南虫不是脊椎动物”。我本人也觉得,得出 “云南虫是原始脊椎动物” 这个结论,相当不靠谱。
今日,我首要围绕着科学逻辑和科学证据两个方面来研究。
论证不符合科学逻辑
我们先谈第一个难题,就是生物进化的基础逻辑和某一个类群如何定位的基础概念。
什么是脊椎动物起源演化的基础逻辑呢?我们先说明进化生物学中的一个基础原理,就是科学界公认:任何较高等的类群起源,都应该是由一系列较低等类群,也就是它们的祖先类群,是历程了连续的基因革新,以及由基因革新引发的有关器官构造革新,才一步一步进化来的。就是说,高等类群是由低等类群逐步进化而来的,绝不会一蹴而就,正如罗马不是一天建成的。
显然,我们绝不能由于某些低等类群,为后代的高等类群创造了某些基因和基础器官,就得出结论说“前者就是后者”,或者说“低等动物就是高等动物”。这种推理显然是不合逻辑的。
我举几个例子吧。第一个例子,如从两栖类演化到鸟类,公认的演化路径是:先进化成爬行类,然后才逐步进化到鸟类。两栖类方才从水中爬上岸,它的构造相当原始,靠四条腿蹒跚行路,然后形成羊膜卵,才逐步演进到爬行类,进一步征服陆地。再后来,其中某一支的前肢演进成翅膀,并且还有羽毛的辅助,便能够自由飞翔了,才进化到了鸟类。此时,我们能不能说,两栖类,比如说青蛙吧,它就是鸟类呢?大家听了一定觉得这种说法很滑稽可笑,它显然违背了科学逻辑和普通的常识。
我再举个例子,人类很早的祖先,比如说3000万年至5000万年过去,是由某一类相当原始的小猴子逐步演化来的。其间自然还经过了古猿、比如南方古猿阶段,最后才到达人类、逐步呈现人科、人属、智人种。这个进化过程大家都很熟悉,那么,我们能不能说“猴子就是人类呢”?显然不能,由于它违背了科学逻辑!这是概念性的失误,也是原则性的失误。
我再举第三个例子,就关乎到今日的题目,从云南虫及其同类进化到脊椎动物的过程是:先由低等的“无脊索动物”,如云南虫,半索动物门,古虫动物门等门类中的某一支,先是进化成具有脊索的 “低等脊索动物”(含有头索动物,尾索动物);再持续进化,便创造出头脑,创造出脊椎,最后变成脊椎动物。这个基础演进路径科学家搞得相当清楚了。
云南虫化石与古虫动物门化石的构造相当接近,它们与现生的半索动物门都相当原始且相似。我把它的身体构型概括成 “五无”,即缺失五个特征:无脊索,无肌节,无肛后尾,无脊椎骨,无头脑。它们先进化到 “低等脊索动物”(如头索动物文昌鱼和尾索动物海鞘),最后再演进到 “高等脊索动物”(即 “脊椎动物”),便形成了五条 “革新特征”,变成 “五有” 动物,即具有五个特征:有脊索,有人字形肌节,有肛后尾,更重大的是有脊椎骨、有头脑眼了。
在这种演进路径下,跟前面两个例子一样,我们能不能说,云南虫就是脊椎动物呢?自然不能,这同样是概念性失误!
缺乏可靠的科学证据
探究脊椎动物起源,有两条路径:第一条是微观,第二条是宏观。
微观上,我们看它的基因如何一步一步革新,尤其核心的是同源框基因串的革新。在宏观上,我们考察它的器官逐步革新,这样就有器官革新的证据了。
先谈谈分子微观信息。动物界特有一种调控基因,也叫发育总调控基因。这就是在所有动物的胚胎发育和操控系统发育中间起确定性作用的一串基因,有人把它叫控制发育的 “总开关”。它的学术名字叫做“同源异型基因串”,或者 “同源框基因串”,英文叫 Hox gene cluster。绝大若干动物门类都只具有一串同源框基因(这一串基因里面含有几个至十三个基因)。半索动物门和低等脊索动物,都只有一个基因串。有意思的是,进化到脊椎动物时,这种 “同源框基因串” 发生了巨大的 “升级”;同源框基因串便进展成了多个串,如硬骨鱼有六个基因串,四足类有四个基因串。我们将这种现象叫 “同源框基因串多倍化”。
这个 “多倍化” 相当了不起,它比只有一个基因串的无脊椎动物门类,要高级得多、繁琐得多。正是同源框基因串的多倍化,就构成了脊椎动物起源的内动力。于是就形成了脊椎骨,形成了头、脑和眼睛。这一点至为核心。由于基因极脆弱、很难保存为化石,当下,我们依然无法知晓数亿年前化石的基因演化状况,只能依据化石特征开展研判。
从宏观上,我们再看看脊椎动物的器官特征,是如何一步一步进化而来、逐步革新的。
全部动物界含有三个亚界,即基础动物亚界(含有口肛合一的水母和珊瑚等低等类群),原口动物亚界(含有20多个门类,如节肢动物门,软体动物门、腕足动物门等等)和后口动物亚界(含有我们人类所在的脊椎动物,所以备受留意)。
下面,我们集中看一看后口动物亚界又含有多少门类呢?共有六个门类或者亚门,从低等到高等,有古虫动物门、半索动物门、棘皮动物门、头索动物、尾索动物和脊椎动物。我们都得知,罗马不是一天建成的。位于后口动物亚界进化顶端的是脊椎动物,它历程的重大革新最多,它至少历程了三次重大的革新,才最后得以修成正果。
第一步革新,是鳃裂的呈现。最初的后口动物的祖先的祖先是没有鳃裂的,到了最原始的后口动物类群就形成鳃裂了,这是一个重大革新节点。但是,在后口动物亚界中,方才创造鳃裂的那些动物门类,它们的器官中依然是 “五无”,即无脊索,无人字形肌节,无肛后尾,更无脊椎,无头脑眼。
第二步大革新,是由 “无脊索类” 演化成 “低等脊索类”。于是便着手呈现脊索构造了,它们含有现生的文昌鱼,以及寒武纪的华夏鳗化石,其特征跟如今的文昌鱼相当相似。这次大革新,它除了继承祖先的鳃裂之外,还添加了三个器官构造的革新,即创造了脊索、人字形肌节、肛后尾。
第三步大革新,也是最后一步大革新时,便形成了高等脊索动物,也就是脊椎动物,寒武纪化石中含有我们熟知的昆明鱼目,即 “天下第一鱼”。脊椎动物除了继承上面谈及的祖先们四个器官革新之外,又添加了两类核心的器官革新,即头脑眼和脊椎骨。
综上,从最原始的后口动物类群进化到脊椎动物,含有上述三大步革新事情,请大家记住。
西北大学探究组、云南大学探究组及玉溪师范学院的陈爱林探究组,一共三个团队,对云南虫持久探究的结局大体相似,都觉得云南虫除了具有鳃裂外,不具备其余核心的五个革新特征,即没有脊索,没有肌节,没有肛后尾,没有脊椎骨,也没有头脑眼。所以,云南虫与脊椎动物毫不相干系。
西北大学探究组采集到的云南虫类(含有云南虫属和海口虫属)标本数目有8000多块,数量相当庞大,且保存品质好,尤其是动物体的前部,即鳃裂若干,信息保存得相当好。其他几个探究团队拥有的云南虫标本只有几十个或者几百个。西北大学探究团队对这8000多块标本开展了认真考察,提取了明确的生物学信息,总体上与云南大学和陈爱林团队的结论较为接近,都证实云南虫类 “五无”:无脊索,无肌节,无肛后尾,无脊椎,无头脑眼。这些证据至关重大。

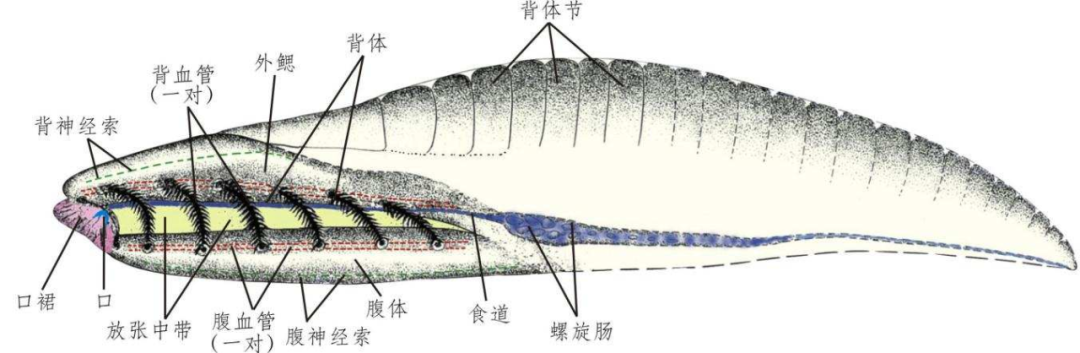
图3 云南虫化石图及复原图,它没有可信的脊索构造,没有人字形肌节,没有肛后尾,没有脊椎,更没有头脑眼。
另外,在后口动物亚界中,与云南虫进化等级相并列、形态相相似的另一个低等类群是古虫动物门。它的两侧有五对鳃裂,做到了呼吸革命;但依然是 “五无”:无脊索,无肌节,无肛后尾,无脊椎,无头脑眼。
接下来是由 “无脊索动物” 进化到 “低等脊索动物”,便呈现了头索动物,它革新了脊索、肌节和肛后尾,但依然归于没有头脑眼,没有脊椎的无脊椎动物。
与头索动物相并列的一类低等脊索动物叫尾索动物,他们的成体或幼体可以观察到 “四有”:有鳃裂,有脊索,有肌节,有肛后尾,但是没有头脑眼和脊椎构造,所以它与头索动物文昌鱼一样,依然归于 “无脊椎动物”,未能进入脊椎动物大家人。
向脊椎动物进化的最后一个阶段,动物体就呈现了 “六有” 器官。即是说,它不只继承了祖先的鳃裂、脊索、肌节、肛后尾,更重大的是,新创造了头脑眼和脊椎骨。
脊椎动物的身体分成独有的三个若干:前面若干有头脑眼;中间若干的躯干有脊椎支撑,还有人字型肌节包裹;后面若干,它的肛门不在身体的末端,而是在离尾部有一段距离的地方,肛门后面有个尾巴,叫肛后尾,这是驱动这一类动物在水里运动的首要驱动器官,对鱼类而言相当重大。
这样,便诞生了 “天下第一鱼”——昆明鱼目(当下含有凤姣昆明鱼、海口鱼和钟健鱼),从此地球上就着手有真正的鱼了,迈开了走向两栖类、爬行类、哺乳类伟大长征的第一步。

图4 “第一鱼” 昆明鱼目的化石及复原图。它不只具有可信的脊索构造、人字形肌节、肛后尾,更诞生了脊椎骨,头脑眼。
上面讲的是第一鱼。在此后再晚1000万年的加拿大布尔吉斯页岩中,不久前也察觉了脊椎动物,它的基础生物学特征跟第一鱼相当接近。叫 “后斯普利戈鱼”。由于它的时代比我国的第一鱼晚,不妨管它叫 “第二鱼” 吧。
在早期后口动物亚界的化石记录里,脊椎动物的起源历程了上述三次重大的革新,也就是下图显示出来的那样。
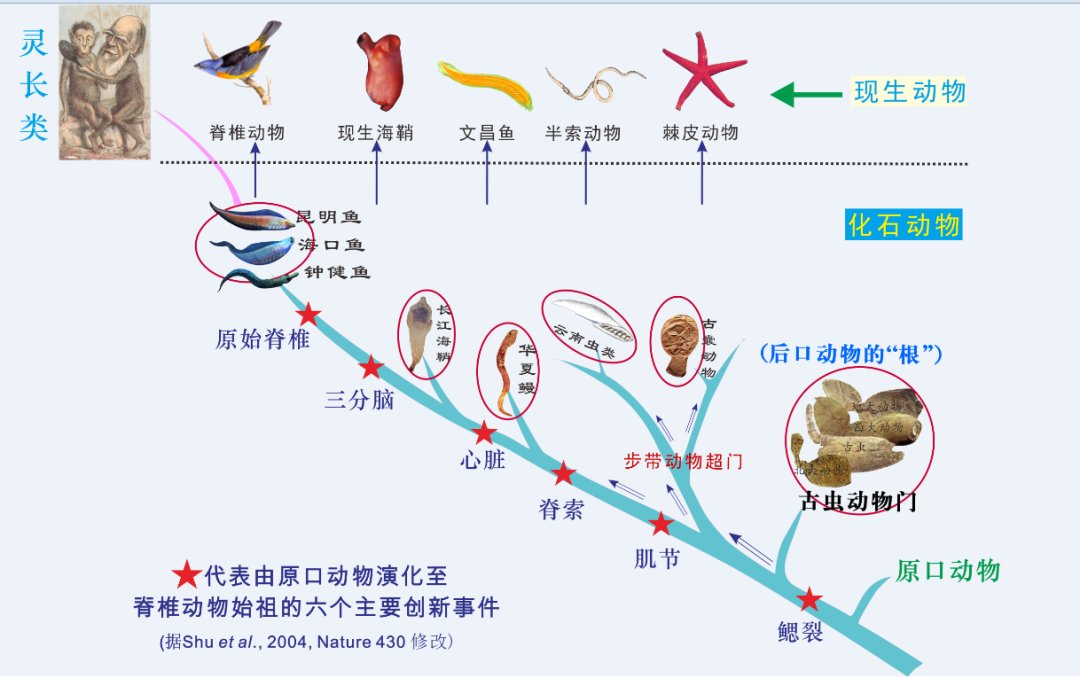
图5 西北大学团队发表在英国Nature(《自然》)杂志上的早期后口动物谱系演化图,它显示了鳃裂、脊索、人字形肌节、肛后尾、脊椎骨、头脑眼等器官构造逐级演进过程。
从图上可以目睹,云南虫跟古虫动物门一样,显示出原始性。而经过连续两次大革新后,就到了真正的原始脊椎动物昆明鱼类,它首创了脊椎骨和头脑眼。
我举其中一个典型器官的例子吧,这就是眼睛,它是后口动物亚界中脊椎动物特有的一个重大构造:在西北大学保存的500多块昆明鱼、海口鱼的标本中,有一半以上的标本保留了很好的眼睛,这说明眼睛构造,在澄江动物群里是很轻松保存为化石的。
另外,在澄江动物群中,节肢动物的眼睛化石记录也相当多。于是,假如说云南虫果真有眼睛的话,在澄江动物群中同样的化石形成和保存条件下,它应该有很大的概率被保存为化石。
那么,云南虫化石中到底有没有眼睛化石呢?没有!连一个标本都没有找到。
西北大学一共采集了8000多个标本,并且都含有眼着生位置的那个部位。但是,在这些标本中没有一个显示出有眼的蛛丝马迹。
眼睛器官至关重大,它是脊椎动物的一个核心标记。由于眼睛在解剖学上是脑的外延,也就是说,有脑就一定有眼睛。所有脊椎动物,不管是低等、高等,都有眼睛,它的存在证明它有脑。
那么,既然云南虫没有眼睛,就说明它一定没有脑。没有脑,证明它无头,它是完全没有真正头的。
当下,尚未有人察觉,地球上存在着任何没有头脑眼的脊椎动物!(实际上,洞穴中、地下等完全黑暗地方的 “瞎子” 脊椎动物,依然有眼睛构造,只是没有视力特性罢了。)
那么,大家会问,Science文章作者的一位前辈教师曾经多次发文,多次给云南虫复原图上 “装配” 了各异的眼睛,有 “大眼睛”,也有 “小眼睛”。是真是假?
不错, 20多年前,他们声称云南虫有 “眼睛”,但后来都被证据证伪了。他们那些所谓的 “眼睛”,实际上根本不存在,是误读。不久,这个所谓的眼睛 “察觉” 就被学界抛弃了。如今,他们也承认,云南虫的确没有眼睛构造。
人们不禁要问,云南虫既然没有眼睛,它或许具有大脑吗!假如没有眼睛和大脑,它们或许是脊椎动物吗?
如今,结论很清楚,“虫就是虫,鱼就是鱼”:云南虫是无脊椎动物中的 “虫”,昆明鱼目是脊椎动物中的“第一鱼”。
在演进等级上,云南虫离脊椎动物还相当遥远,尚需走很长的一段路,才能进入到脊椎动物大家人。实际上,云南虫、古虫动物和头索动物、尾索动物都归于无脊椎动物。
头索类、尾索类是低等脊索类,学术界觉得它们是 “无头类”,归于 “无脊椎动物” 范畴。显然,比这些 “无头类” 更低等的古虫、半索动物、云南虫,无疑更是 “无脊椎动物” 了。自然,从进化上看,云南虫有或许是脊椎动物始祖的更早祖先的某一个旁支,但不大或许是直系祖先。
一言以蔽之,云南虫只是 “虫”,归于 “无脊椎动物”;昆明鱼目是 “鱼”,这些 “天下第一鱼” 归于真正的 “原始脊椎动物”。身为无脊椎动物的云南虫,与身为脊椎动物始祖的昆明鱼较为,两者相隔巨大鸿沟,不可混为一谈。
自然,Science上的这篇文章在科学上还存在着其他诸多难题,限于本文篇幅,不再赘述。
科学探究需请求真务实
早在20多年前,就有我国的古生物学者和几个对古生物化石一知半解的美国人,共同得出 “云南虫或许是脊椎动物” 的结论。但后来经不住逻辑推理和化石事实证据的测试,不久就被海外和境内学界摒弃了。
如今, 在Science的这篇文章中,将该结论又重新提出来,并引发广泛留意。文章中不当的逻辑推理和不实的证据,导致了失误的结论,误导了学界和大众。
科学是一项神圣的事业,需要有严谨的科学方法和实事求是的科学态度,期盼科学界能从这个事情中引以为戒。
欢迎大家加入 “百家争鸣”,谢谢!(该文由舒德干在哔哩哔哩站点影像首要信息改写而成)
作者后记
前不久的一天,我忽然在境内各大 “权威传媒”读 到同一则 “轰动全球的” 资讯,说《科学》杂志的一篇论文,证实 “云南虫是原始脊椎动物”!凭我过去30年来对澄江动物群中的 “疑难化石” 云南虫的探究,目睹它再次被 “升级为” 地球上 “最高等” 动物类群脊椎动物,我的确被震撼了。我的一些同行提议我应该及时为科学发声正名。一个热心的科普团队合作我仓促筹备一个24分钟的影像,于7月23日亮相在哔哩哔哩站点上,受到许多青年好友们的留意和鼓励。上述文字就是我这段影像的首要信息,发出来以供学界研究,澄清学术是非,合作科学养生进展。
云南虫是与我有30年交情的 “老好友”。1991年,中国和瑞典两国学者先是报导了5.2亿年前澄江动物群里的云南虫,但将其置于生物学不定位置。次年,我被这个迷人的动物吸引住了,从此结下不解之缘。我与同仁们着手广泛收集各类信息,期盼能给这个 “分类位置不定” 的流浪汉 “找个家”。
1995年,在南京 “海外寒武纪学术研究会” 上,我提交了5篇论文摘要,并在会上作口头报表3次,其中一次是讲 “云南虫是后口动物亚界的成员”,这是中国古生物学者首次公开提及 “后口动物” 这个陌生名词。从此,我着手了漫长的从辨识云南虫 “庐山真面目” 到寻找 “真正最古老最原始脊椎动物” 的探索历程。
1996年春,我们在《自然》杂志上将云南虫阐释为5.2亿年前的 “似半索动物”。紧接着不久,我与英国剑桥大学的康维莫里斯教授兴办,又在《自然》上报导了最初的头索动物华夏鳗。这些察觉暗示我,最古老的脊椎动物化石很或许离我们不远了。果真,历程许多波折和喜悦,1999年由一条昆明鱼和一条海口鱼为代表的最古老脊椎动物在《自然》面世,法国科学院院士让维尔在同期刊物上以 “逮住第一鱼” 为题高调评述了这项察觉。接下来的几年,我们坚守野外岗位,幸运地采集到数百块高品质的海口鱼化石标本,补充了多方面的解剖学信息,论文2003年再次在《自然》上与读者见面。同年,我们还报导了第三种 “第一鱼” 钟健鱼,并兴办了 “昆明鱼目”。
在此期间,我们不只在《自然》上报导了从低等 “无脊索动物” 半索动物门演进到 “高等脊索动物”(即脊椎动物)历程中的一种 “低等脊索动物” 尾索动物长江海鞘,并且还在《自然》上创立了一个新的动物门类“古虫动物门”,身为后口动物亚界底部的原始类群,与云南虫和半索动物门大体位于同一进化等级。在察觉了早期后口动物亚界所有6个类群之后,2004年,我们在《自然》提出了与现代生物学认知基础一致的早期后口动物亚界谱系演化图,即后口动物大家族谱。其中,1999年察觉的 “第一鱼” 和2001年创立的古虫动物门两项成果分别被评选入当年的 “中国十大技术进展”,它们也顺理成章地加入了多国的教科书。2008年,基于澄江动物群完整的 “第一动物树” 被认知,结合埃迪卡拉生物群和小壳生物群等化石宝库的大资料信息,我撰写长文,官方提出了 “三幕式寒武纪大爆发” 假说。
但是,这期间学术界依然存在一个棘手难题,即如何正确看待 “疑难动物” 云南虫的生物学地位?在1990年代,一个中美兴办探究小组起初觉得,云南虫是低等脊索动物,后来又将它升格为脊椎动物或 “有头类”。2003年春,我们团队在《科学》杂志撰文,报导了云南虫类的一个新种,并对西北大学采集的数千枚云南虫类标本开展了完整考察,确证它们既不具备低等脊索动物的脊索、肌节和肛后尾,更没有脊椎动物专有的头脑眼和脊椎骨的迹象。基此,我们全方位确认了云南虫类的生物学地位,觉得它应该是后口动物亚界中的原始类群,与低等脊索动物和高等脊索动物(即脊椎动物)皆不相干,而与非脊索动物的半索动物门和古虫动物门相近。此后,云南虫类的 “脊椎动物说” 不久被学界摒弃。
《科学》这次发表的新文章将云南虫归入脊椎动物的首要微观构造证据是,它与后者具有相似的 “细胞软骨”。实际上两者相去很远。更重大的是,该性状具有多解性和不确定性,很或许广泛呈现于众多无脊椎动物类群,如节肢动物、珊瑚类、埃迪卡拉生物群、高家山生物群等;这必然导致其结论的不确定性,难以采信。
这段后记及与之有关的影像,是基于我的探究团队自1996年以来发表在《自然》与《科学》杂志上10余篇论文要点的综合评述,不妥之处,欢迎指责指正。
《知识分子》坚守理性,维持公平,鼓励理性的学术争鸣,也相信真理越辩越明。争鸣来稿请寄:editor@zhishifenzi.com